29 papers mentioning mmu-mir-215
Open access articles that are associated with the species Mus musculus
and mention the gene name mir-215.
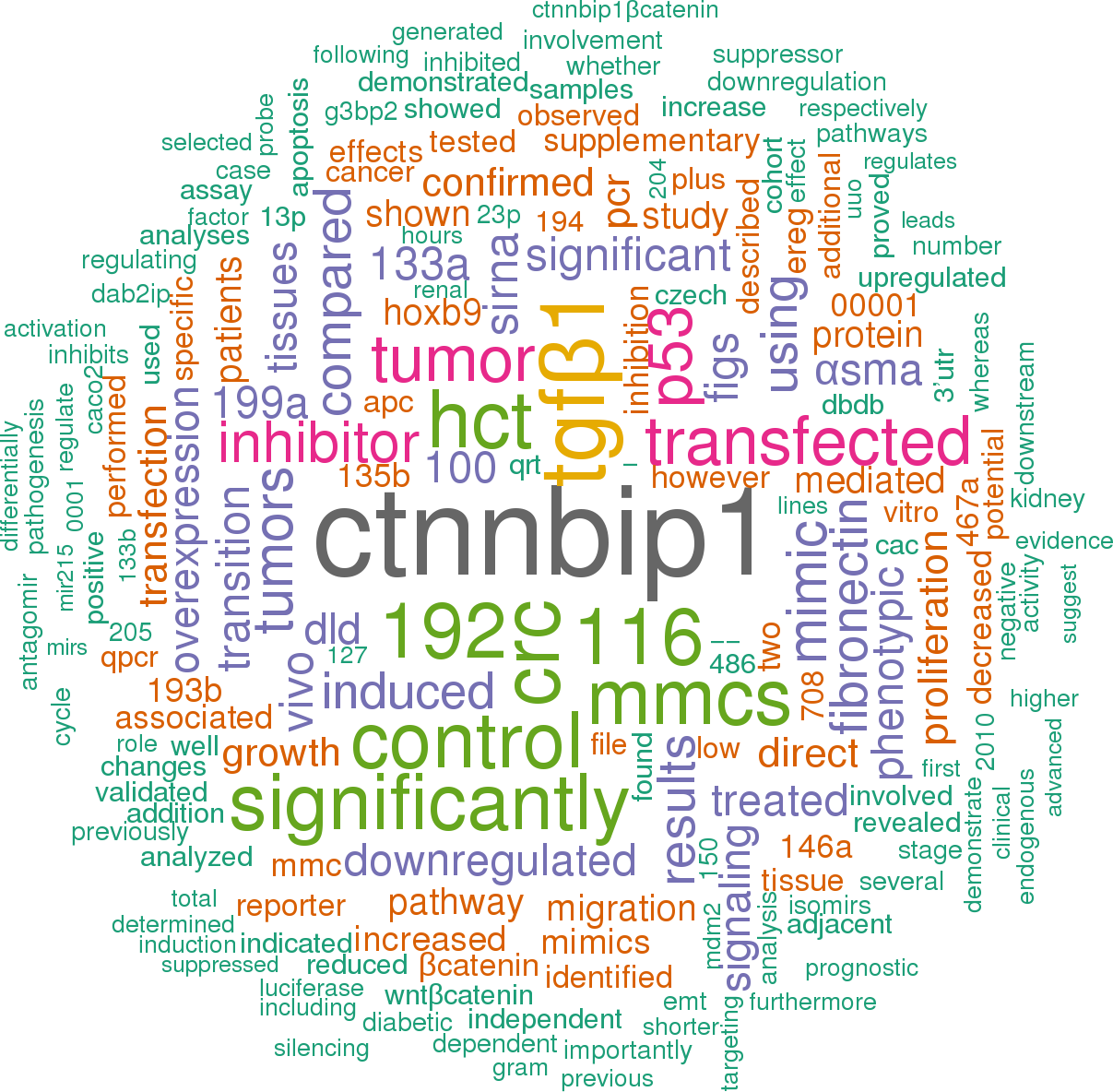
Click the buttons to view sentences that include the gene name, or the word cloud on the right for a summary.
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|